
IF:27.4 《AM》南京师范大学万密密、毛春:人工线粒体纳米机器人通过口服在体内输送能量
专栏:学术前沿
发布日期:2025-04-30
缺血性心脏病(IHD) 是全球首要死亡原因,其核心病理特征为心肌细胞线粒体功能障碍,导致能量代谢紊乱。因此,开发增强线粒体 ATP 产生能力的治疗方法对治疗此类疾病具有重要意义。
目前,增强细胞 ATP 产生能力的方法主要有两类:一是直接移植活性线粒体,理论上可完全替代受损线粒体,但因线粒体分离脆弱、纯化保存复杂、免疫相容性等问题,临床应用面临挑战;二是构建人工 ATP 产生系统,虽在体外有效,但大多依赖额外能量输入(如可见光),且穿透深度有限,体内效率受限。目前尚未有稳定、不依赖额外能量输入且可用于体内的成熟人工 ATP 合成系统。

针对上述问题,南京师范大学万密密、毛春等人设计了一种口服人工线粒体纳米机器人(AMNs),用于修复受损细胞的能量供应系统,直接提供ATP。该纳米平台由两性离子单体MAPCr(含运动和能量产生单元)和PFA(屏障穿越单元)制备而成。在IHD模型中,AMNs的PCr部分可与细胞质中的ADP反应生成ATP,修复能量供应系统。其PFA部分可穿透肠黏液屏障进入血流,L-精氨酸部分则靶向受损线粒体。到达受损心脏后,AMNs能快速产生ATP,减轻线粒体负担,恢复细胞活性,同时与ROS反应生成NO,减轻炎症并促进线粒体活性。该文章于2025年4月14日以《Artificial Mitochondrial Nanorobots Deliver Energy In Vivo by Oral Administration》为题发表于《Advanced Materials》(DOI:10.1002/adma.202500495)。
(1)PFMACr AMNs 的制备和表征
选择两性离子N-甲基丙烯酰基-L-精氨酸-磷酸肌酸(MAPCr)和丙烯酸十五烷基氯辛酯(PFA)作为单体制备PFMACr AMN(图1A)。在IHD模型中,AMNs的PCr部分可与细胞质中的ADP反应生成ATP,修复PCr-CK供能系统(图1B)。口服后,PFMACr AMN的PFA部分可穿透肠黏液屏障进入血流(图1C)。TEM结果显示纳米颗粒呈均匀球形(图1D),DLS测量其流体动力学直径约为150 nm(图1E),添加PFA的样品ζ电位降低(图1F)。共聚焦图像和DLS结果表明PFMACr AMN表面无蛋白质粘附(图1G,H)。处理后,损伤心肌细胞中ROS水平恢复正常,NO水平增加至正常心肌细胞的2.8倍(图1I-K),这可能是由于MAPCr组分消耗ROS并产生NO。PFMACr AMNs在10 mM GSH的模拟生理环境中12小时内迅速降解后保持稳定(图1L),表明其可在细胞环境中降解并暴露活性位点。与仅维持3小时ATP合成能力的纯PCr相比,PFMACr AMN可使受损心肌细胞内ATP水平持续增加至少12小时(图1M)。400 μg AMNs在10⁵受损心肌细胞中的ATP生产能力与10⁷天然线粒体相当(图1N)。
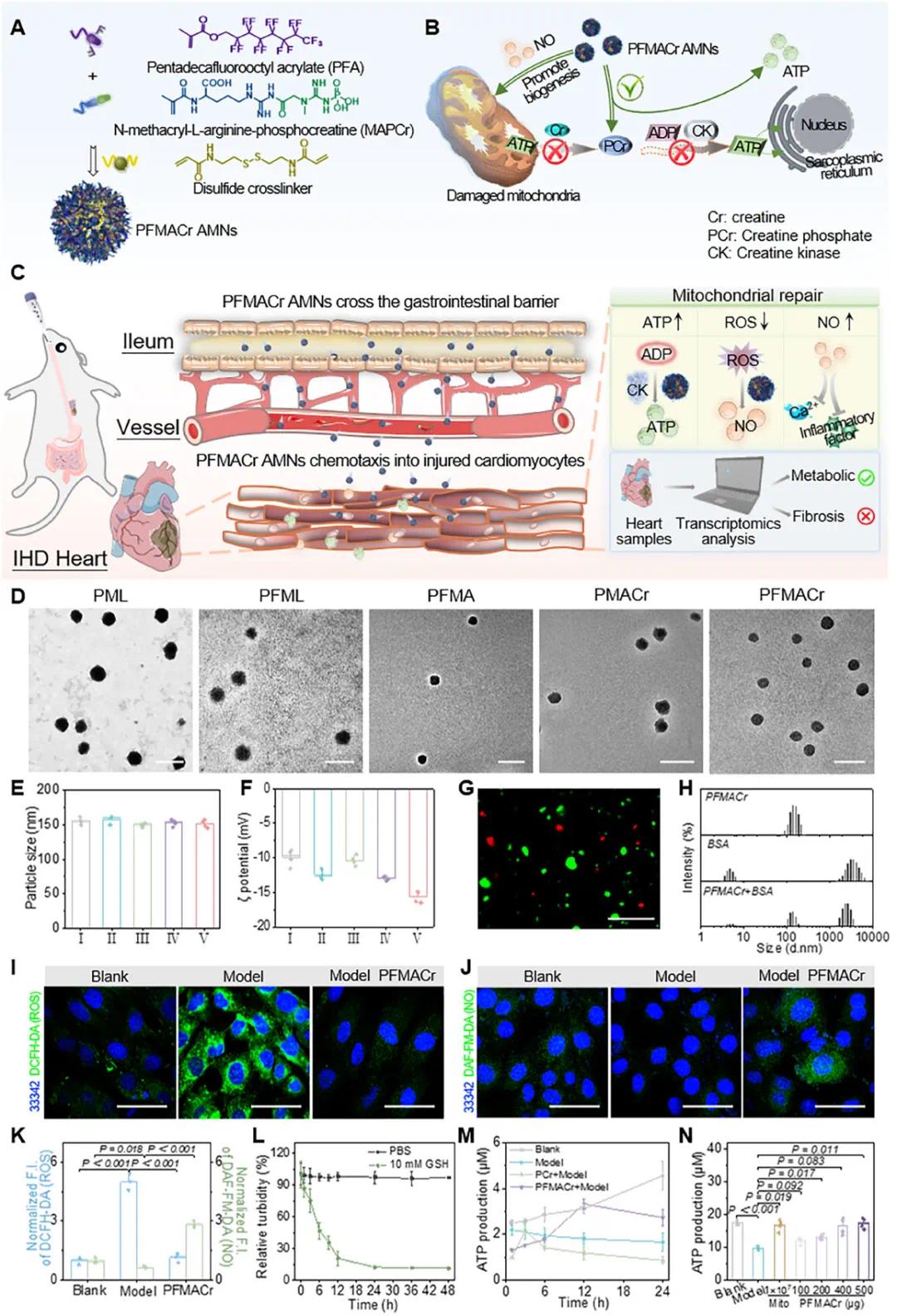
图1 PFMACrAMN的制备、口服给药及基本性质。(a)PFMACrAMN的制备方法;(b)PFMACrAMNs产生NO促进受损线粒体生物合成,并提供PCr促进ATP产生;(c)PFMACrAMN口服给药的体内递送及治疗途径;(d)不同样品的TEM图像(比例尺:200 nm);(e,f)分散在PBS中的样品的水合粒度和电位(I:PML,II:PFML,III:PFMA,IV:PMACr,V:PFMACr);(g)Cy5标记的PFMACrAMN(红色)和FITC标记的BSA(绿色)混合溶液在37℃孵育4 h后的荧光图像(比例尺:10 µm);(h)37℃孵育4 h后PFMACrAMN(1 mg/mL)和BSA(500 mg/mL)的DLS结果;(i,j)PFMACrAMN处理的缺氧损伤H9c2细胞中ROS(i)和NO(j)水平的CLSM图像(比例尺:50 µm);(k)i和j的归一化荧光强度;(l)PFMACrAMNs在PBS或10 mM GSH中降解48 h的相对浊度;(m)PFMACrAMN在缺氧损伤H9c2细胞中产生ATP的能力;(n)PFMACrAMN与天然线粒体在缺氧损伤H9c2细胞中产生ATP能力的比较(n=3-4)
(2)趋化运动行为
PFMACr AMNs中的L-Arg组分能够响应损伤心肌细胞中高表达的iNOS,诱导其在损伤细胞中的趋化运动。在静态Y形微流体通道模型中,正常H9c2细胞环境中,PFML NPs和PFMACr AMNs呈典型布朗运动;而在LPS预刺激损伤的H9c2细胞中,PFML NPs仍保持布朗运动轨迹(速度小于1 µm/s),PFMACr AMNs的运动速度则显著提高至5.2 µm/s(图2A、B)。在静态模型中,将含有正常和预刺激H9c2细胞裂解物的琼脂糖凝胶填充到通道中,PFML NPs在室II和室III的荧光强度无显著差异(图2C、D、E),而PFMACr AMNs在室III的荧光强度显著高于室II,且60分钟时差异最大,室III荧光强度是室II的6.7倍(图2F、G),表明PFMACr AMNs能够感知高浓度iNOS并沿其浓度梯度扩散。在动态“Y”型微流体模型中,当两侧通道均填充正常H9c2细胞裂解物时,PFML NPs和PFMACr AMNs均未表现出趋化行为(图2H、I、J);而当一侧通道填充预刺激H9c2细胞裂解物时,PFMACr AMNs的红色荧光向预刺激侧移动,移动距离为1095.3 µm,而PFML NPs未表现出趋化行为(图2K、L)。
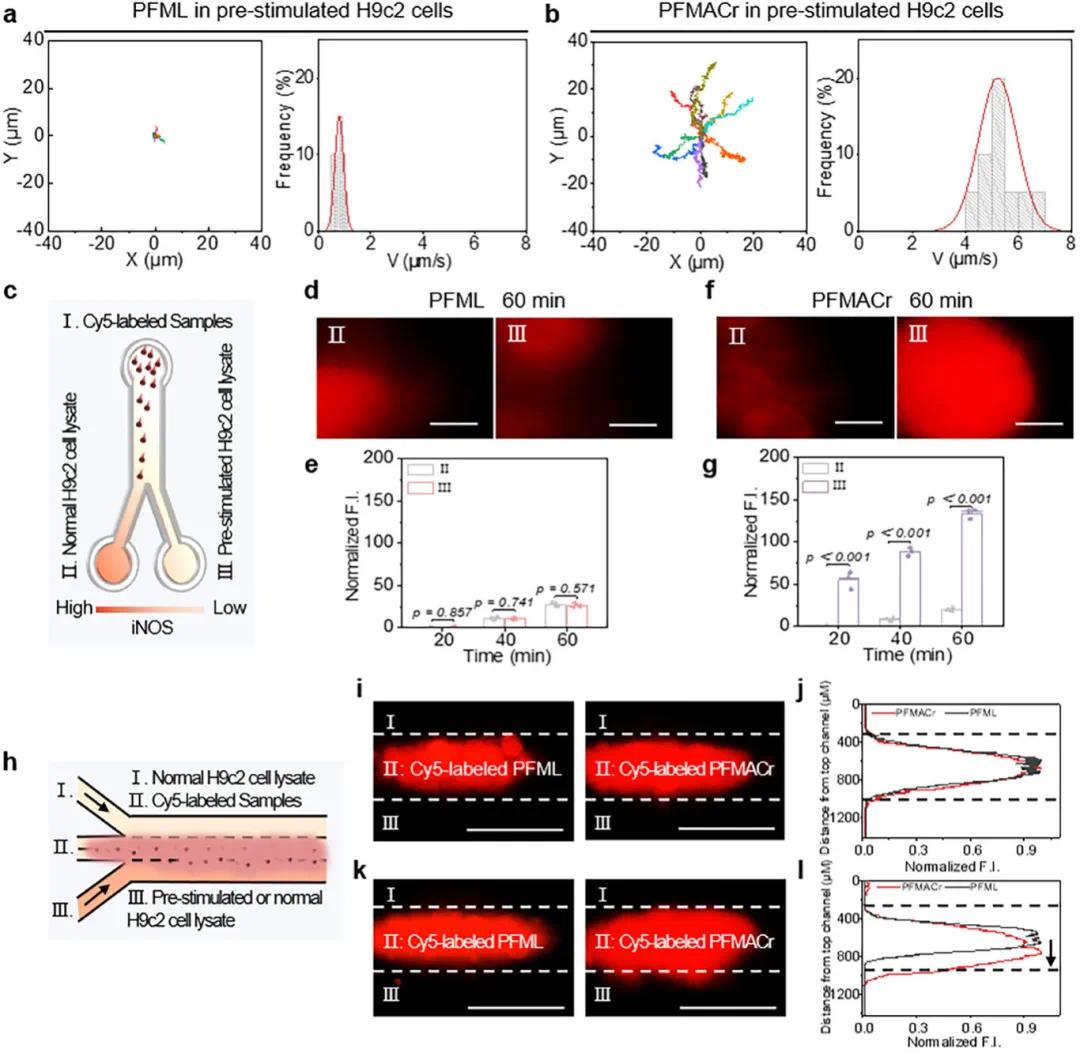
图2 PFMACr AMNs在模拟IHD微环境中的运动行为。(a,b)PFML NP(a)和PFMACr AMN(b)在预刺激H9c2细胞中的轨迹和速度分布(20 s,n=10);(c)静态Y形通道示意图;(d,e)PFML NP添加后不同时间室II和室III的荧光图像(60分钟)(d)及归一化荧光强度(e)(比例尺:500 µm);(f,g)PFMACr AMNs添加后不同时间室II和室IV的荧光图像(60分钟)(f)及归一化荧光强度(g)(比例尺:500 µm);(h)动态微流控装置示意图;(i,j)在两个通道中应用正常H9c2细胞裂解物的微流体装置中PFML NP和PFMACr AMN的代表性荧光图像(i)及标准化荧光强度(j)(比例尺:500 µm);(k,l)施加正常H9c2细胞裂解物和预刺激H9c2细胞裂解物的微流体装置中PFML NP和PFMACr AMN的代表性荧光图像(k)及归一化荧光强度(l)(比例尺:500 µm)
(3)损伤细胞对PFMACr AMNs的细胞摄取行为
PFMACr AMNs能够有效穿透血管内皮细胞屏障并被受损心肌细胞摄取。在2D Transwell模型中,PMACr NM的穿透能力优于PML NP,而PFMACr AMNs的穿透能力最强(图3A-C)。在缺氧损伤的H9c2细胞中,PFMACr AMNs的摄取表现为更突出的红色荧光信号(图3D,E)。摄取抑制剂实验表明,PFMACr AMNs进入心肌细胞主要通过小窝蛋白介导的途径和能量依赖的内吞途径。细胞器定位实验显示,PFMACr AMNs聚集在线粒体附近,可能是因为线粒体损伤时释放大量ROS,而PFMACr AMNs倾向于聚集在ROS浓度高的区域以转化为NO。此外,其修饰有助于通过疏水相互作用与膜脂质融合,并通过疏脂性诱导快速跨膜扩散,增强溶酶体逃逸功能(图3F,G)。
(4)体外治疗效果
PFMACr AMNs的MAPCr可与H₂O₂反应产生NO。缺氧损伤的H9c2细胞中H₂O₂含量是正常细胞的2.3倍(图3H)。PFML NPs处理后H₂O₂未减少,而非氟PMACr NMs和PFMACr AMNs能消耗缺氧细胞中的H₂O₂,使O₂⁻和MDA降至空白组水平(图3I)。氧化应激缓解后,损伤相关分子模式释放减少,促炎基因表达和炎性细胞因子分泌被抑制,TNF-α和IL-6水平回到空白组水平(图3J)。PFMACr AMNs通过NO降低细胞质Ca²⁺水平,缓解线粒体钙超载,经Fluo-4 AM标记后,缺氧细胞经PFMACr AMNs处理的Ca²⁺水平恢复正常(图3K,L)。JC-1标记显示,PMACr NM和PFMACr AMN处理可使线粒体膜电位恢复(图3M,N)。NO还可上调PGC-1α水平,刺激线粒体生物合成(图3O)。PFMACr AMNs参与PCr-CK能量穿梭快速合成ATP,与缺氧损伤的H9c2细胞共孵育后,ATP合成可持续12小时,使缺氧损伤细胞中ATP水平与正常细胞相当(5.6和5.2 µm)(图3P)。此外,PFMACr AMNs还能恢复细胞活力,48小时内可将缺氧损伤的H9c2细胞活性维持在91%(图3Q)。
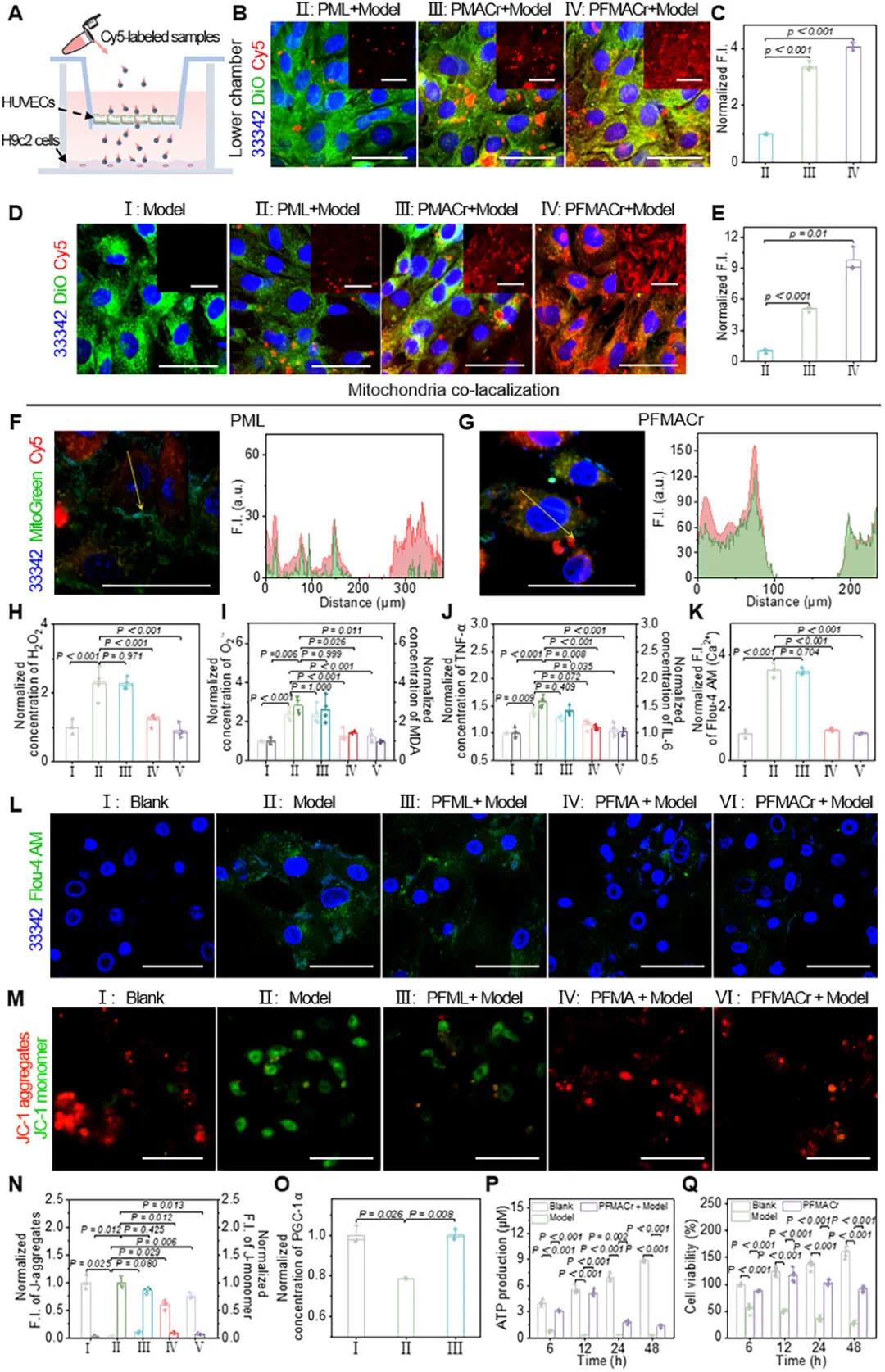
图3 PFMACr AMNs的细胞摄取、选择性胞吐和体外治疗作用。(a)模拟体外HUVEC的PFMACr AMNs渗透Transwell的示意图;(b,c)缺氧损伤的H9c2细胞(Transwell下室)的CLSM图像(b)和归一化荧光强度(c)(比例尺:50 µm);(d,e)缺氧损伤的H9c2细胞中不同样品的摄取CLSM图像(d)和归一化荧光强度(e)(比例尺:50 µm);(f,g)缺氧损伤的H9c2细胞线粒体中PML NP(f)和PFMACr AMN(g)的CLSM图像及共定位结果(比例尺:50 µm);(h-j)不同样品处理后缺氧损伤的H9c2细胞中H₂O₂(h)、O₂⁻和MDA(i)以及炎症水平(j);(k,l)不同样品处理后缺氧损伤的H9c2细胞中Ca²⁺水平和线粒体状态的归一化定量结果(k)及CLSM图像(l)(比例尺:50 µm);(m,n)不同样品处理后缺氧损伤的H9c2细胞线粒体状态的CLSM图像(m)和标准化定量结果(n)(比例尺:50 µm);(o)PFMACr MNs处理后缺氧损伤的H9c2细胞中PGC-1α浓度结果;(p)PFMACr AMNs处理后缺氧损伤的H9c2细胞中ATP水平变化;(q)PFMACr AMNs处理后缺氧损伤的H9c2细胞活性
(5)评价 PFMACr AMNs 在体外和体内突破多重屏障并靶向受损心脏的能力
PFMACr AMNs在胃肠道环境中保持稳定,其粒径在pH 1.2和pH 6.8条件下24小时内不变(图4A、B)。酸碱处理后的PFMACr AMNs与缺氧损伤的H9c2细胞共培养,仍能显著提升细胞ATP水平(图4C),并将NO水平恢复至正常(图4D)。其PFA组分可防止粘蛋白吸附,使其在小鼠肠黏液中的扩散位移和速度高于PMACr NP。Transwell和离体小肠组织实验证明PFMACr AMNs能非破坏性地穿透肠屏障。离体器官成像显示,口服PFMACr AMNs后,其在大鼠肠组织中的荧光强度0.5小时达高峰,3小时内肠系膜微血管荧光强度是PML NPs处理的2.6倍(图4E-I)。
在小鼠皮下肿瘤模型中,PFMACr AMNs静脉注射后10分钟即可突破血管内皮屏障,出现在病变组织中(图4J、K)。在正常小鼠心脏中,PFMACr AMNs主要停留在血管内,而在受伤心脏模型中,其可在口服给药12小时后积聚于心脏组织。与健康大鼠口服PFMACr AMNs(I组)、IHD大鼠口服PML NPs(II组)相比,IHD大鼠静脉注射(III组)和口服(IV组)PFMACr AMNs后,在受损心脏中荧光信号更强(图4N)。口服给药的PFMACr AMNs在IHD损伤心脏中的靶向效率是静脉内给药的16.3%,而静脉内给药的靶向效率是口服给药的135%(图4O,P)。在健康心脏近端区域,III组和IV组PFMACr AMNs的荧光强度分别是I组的3.9倍和2.0倍;在iNOS富集的心脏中部和远端区域,III组PFMACr AMNs的荧光累积分别是I组健康心脏远端区域的10.6倍和8.8倍。研究表明,AMNs在心脏损伤部位的蓄积量大于健康部位,且IV组口服PFMACr AMNs显著增强了其浸润损伤心脏的能力(图4Q,R)。
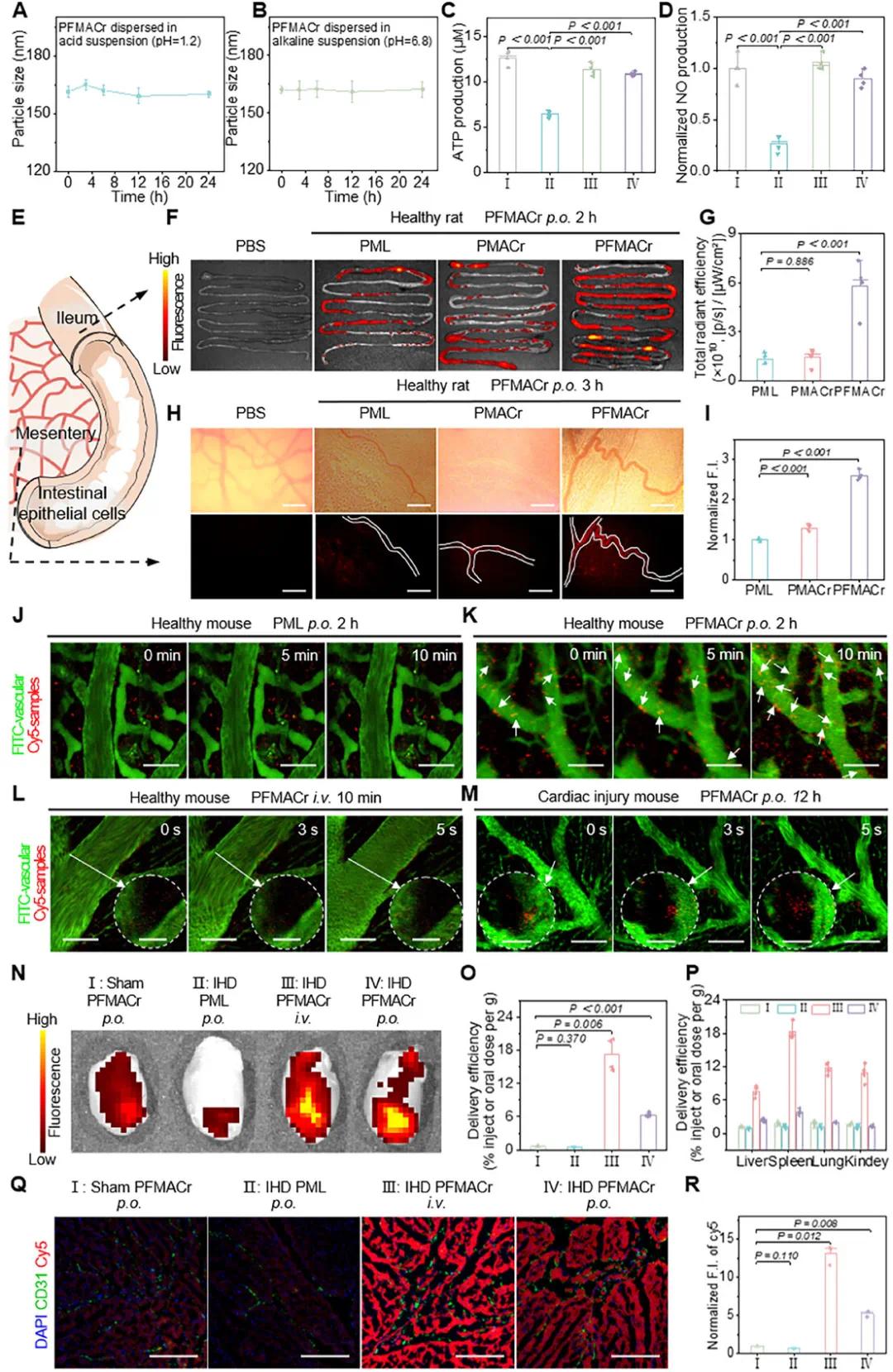
图4 PFMACr AMN的稳定性及其跨越多种生理屏障靶向缺血心脏的能力的表征。(a,b)PFMACr AMNs在pH 1.2(胃pH)(a)和pH 6.8(肠pH)(b)环境中的DLS监测结果;(c,d)经酸(2小时)-碱(6小时)处理的PFMACr AMNs在缺氧损伤的H9c2细胞中产生ATP(c)和NO(d)的能力,对照为未处理的PFMACr AMNs(I:空白,II:模型,III:未处理的PFMACr AMN,IV:处理的PFMACr AMN);(e)样品穿过肠屏障的观察点示意图;(f,g)口服施用不同cy5标记样品2小时后大鼠肠组织的离体图像(f)和定量结果(g);(h,i)口服施用不同cy5标记样品3小时后大鼠肠系膜微血管系统的图像(h)和归一化荧光强度(i)(比例尺:100 µm);(j,k)健康C57BL/6小鼠口服施用PML NP(j)和PFMACr AMN(k)2小时后,基于共聚焦技术的体内显微镜捕获的小肠部位连续图像(比例尺:100 µm);(l,m)健康C57BL/6小鼠静脉内注射PFMACr AMNs(l)后10分钟和心脏损伤C57BL/6小鼠口服施用PFMACr AMNs(m)后12小时,基于共聚焦技术的体内显微镜捕获的从血管释放到心脏组织中的PFMACr AMNs的连续图像(比例尺:100 µm);(n-p)大鼠口服或静脉内施用12小时后心脏(n)和向心脏(o)及其他主要器官(p)的递送效率的离体图像;(q,r)大鼠经口或静脉给药12小时后cy5标记样品在心脏梗死区分布的免疫荧光图像(q)和相应的归一化荧光强度(r)(比例尺:200 µm)
(6)体内治疗效果
在心肌梗死(MI)模型中研究PFMACr AMNs的治疗效果(图5A)。第7天,静脉内施用的PFMACr AMNs(EF:87.7%)比口服的(EF:78.9%)更好地恢复心脏功能(图5B-F),与靶向效率差异一致(图4O)。增加口服给药频率(第VII组)后,IHD大鼠心脏功能持续改善,第7天EF达79.7%,但仍低于静脉给药组。静脉注射组心脏功能在给药周期结束后逐渐恶化,而持续口服组在整个28天期间保持稳定,表明增加口服频率可阻止心脏功能恶化。PFMACr AMNs的疗效优于临床药物BB(III组),可能因其直接合成ATP,优化缺血心肌的能量代谢。给药28天后,通过TTC染色观察各组大鼠心脏损伤面积(图5G),结果显示,与对照组(38.8%)相比,PFMACr AMNs口服7天组(V组)、静脉注射7天组(VI组)和口服28天组(VII组)均有效缩小梗死面积,左心室AOI比例分别降至14.9%、8.7%和10.5%(图5H)。Masson染色(图5I、J)显示,V组、VI组和VII组纤维化程度显著低于对照组和BB组。免疫组织化学CD31染色(图5K、L)显示,PFMACr AMNs治疗显著增加CD31阳性细胞数量(V组:11.8%,VI组:16.6%,VII组:15.9%),改善缺氧损伤区域血供。长期口服PFMACr AMNs能有效预防疾病进展,但前7天治疗效果低于静脉给药组(EF:87.7%)。为验证是否与累积量差异有关,增加单次口服剂量(从10 mg/kg增至50 mg/kg),心脏靶向效率从6.3%变化至5.0%,绝对累积量从126 µg/g增加至500 µg/g(图5M、N)。增加剂量后,第7天IHD大鼠心脏功能恢复(EF:85.9%)接近静脉注射组,并在第28天保持稳定,接近健康大鼠功能(图5O、P)。
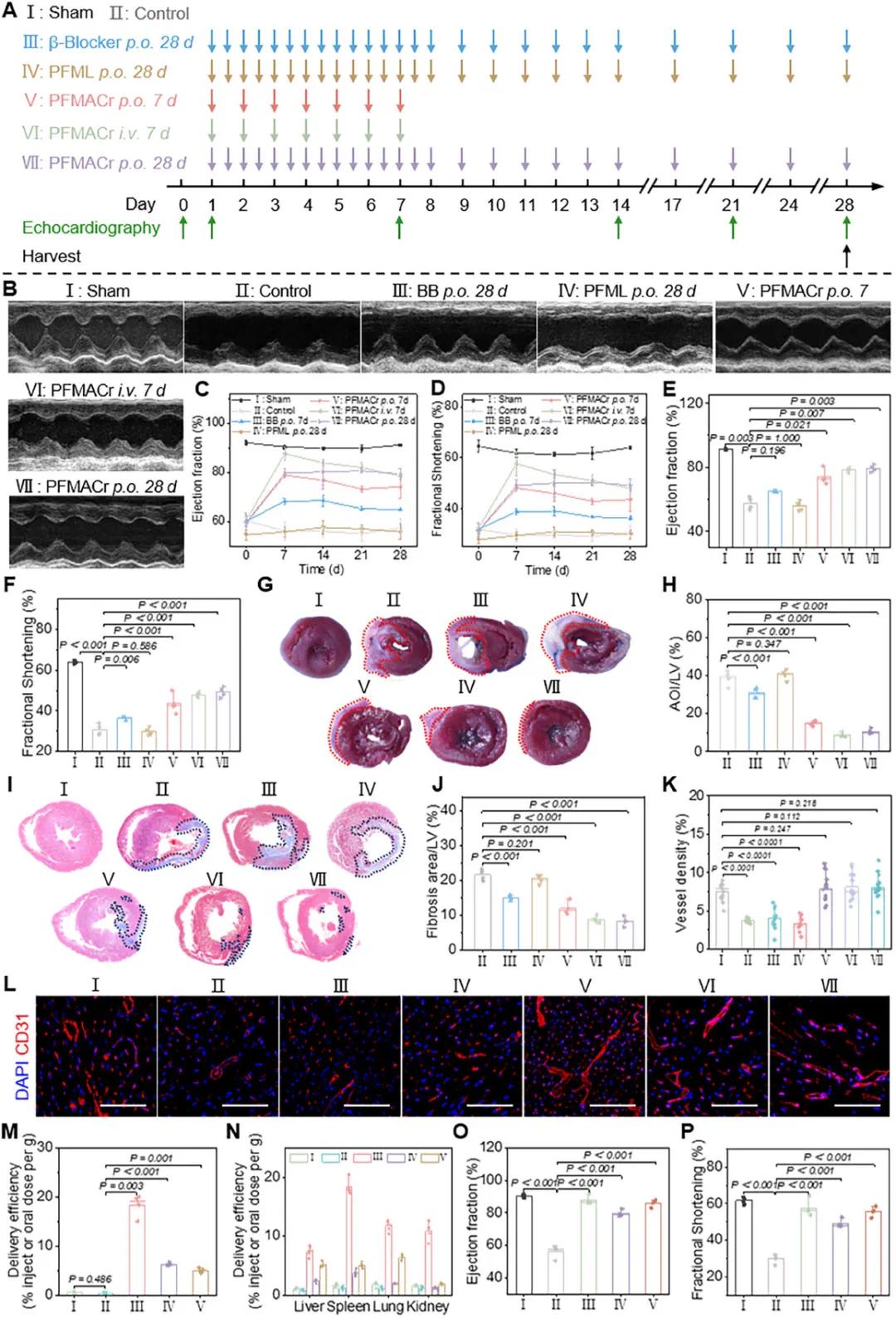
图5 PFMACr AMNs对缺血性心脏病大鼠的治疗作用。(a)IHD大鼠治疗方案示意图,未治疗的IHD大鼠作为对照组;(b)治疗后IHD大鼠的M型超声心动图;(c,d)治疗期间IHD大鼠的EF(c)和FS(d)曲线;(e,f)治疗后IHD大鼠中EF(e)和FS(f)的定量结果;(g,h)梗塞区域以虚线圈出的代表性图像(g)及IHD大鼠TTC染色心脏切片中LV的AOI定量(h);(i,j)纤维化区域以虚线圈出的代表性图像(i)及IHD大鼠Masson染色心脏切片中LV纤维化区域的定量(j);(k,l)IHD大鼠CD31免疫染色心脏切片的血管密度(k)和CLSM图像(l)的定量(比例尺:100 µm);(m,n)大鼠口服或静脉内施用12小时后向心脏(m)和其他主要器官(n)的递送效率;(o,p)PFMACr AMNs治疗7天后大鼠EF(o)和FS(p)的定量结果(前7天每天两次,第8-14天每天一次,第15-28天每周两次);(I:假手术大鼠;II:未处理的IHD大鼠;III:IHD大鼠静脉内给予PFMACr AMN,每次10 mg/kg bw;IV:IHD大鼠口服给予PFMACr AMN,每次10 mg/kg bw;V:IHD大鼠口服给予PFMACr AMN,每次50 mg/kg bw)
(7)治疗后心脏组织的心脏多组学分析
对自然线粒体组(Mito组)和AMNs治疗组(PFMACr组)的大鼠心脏进行转录组分析。基于GO和KEGG富集分析(图6A),天然线粒体移植显著上调与线粒体代谢和结构功能相关的基因,包括线粒体膜、基质、内膜、脂肪酸β-氧化、TCA循环和NADH代谢过程。AMNs治疗组的基因测序结果也显示类似上调基因(图6B),表明AMNs可在基因水平上发挥与天然线粒体类似的功能。同时,下调基因分析显示,天然线粒体处理后,与细胞凋亡、伤口愈合和缺氧反应相关的基因显著下调(图6C),表明心肌细胞活力和适应性增加;AMNs处理后,与炎症、凋亡和纤维化相关的基因也显著下调(图6D),表明AMNs修复了心脏功能。天然和AMNs治疗后心脏受损区域的基因分析显示,两者之间仅有262个DEG,且PFMACr对炎症和免疫相关通路表现出调节作用(图6E、F),包括上调白细胞介素-27信号通路和下调免疫球蛋白介导的免疫应答。基因集富集分析(GSEA)表明,与对照组相比,Mito组和PFMACr组中与氧化磷酸化和TCA循环相关的基因表达上调,而与细胞纤维化相关的基因表达被抑制(图6G)。
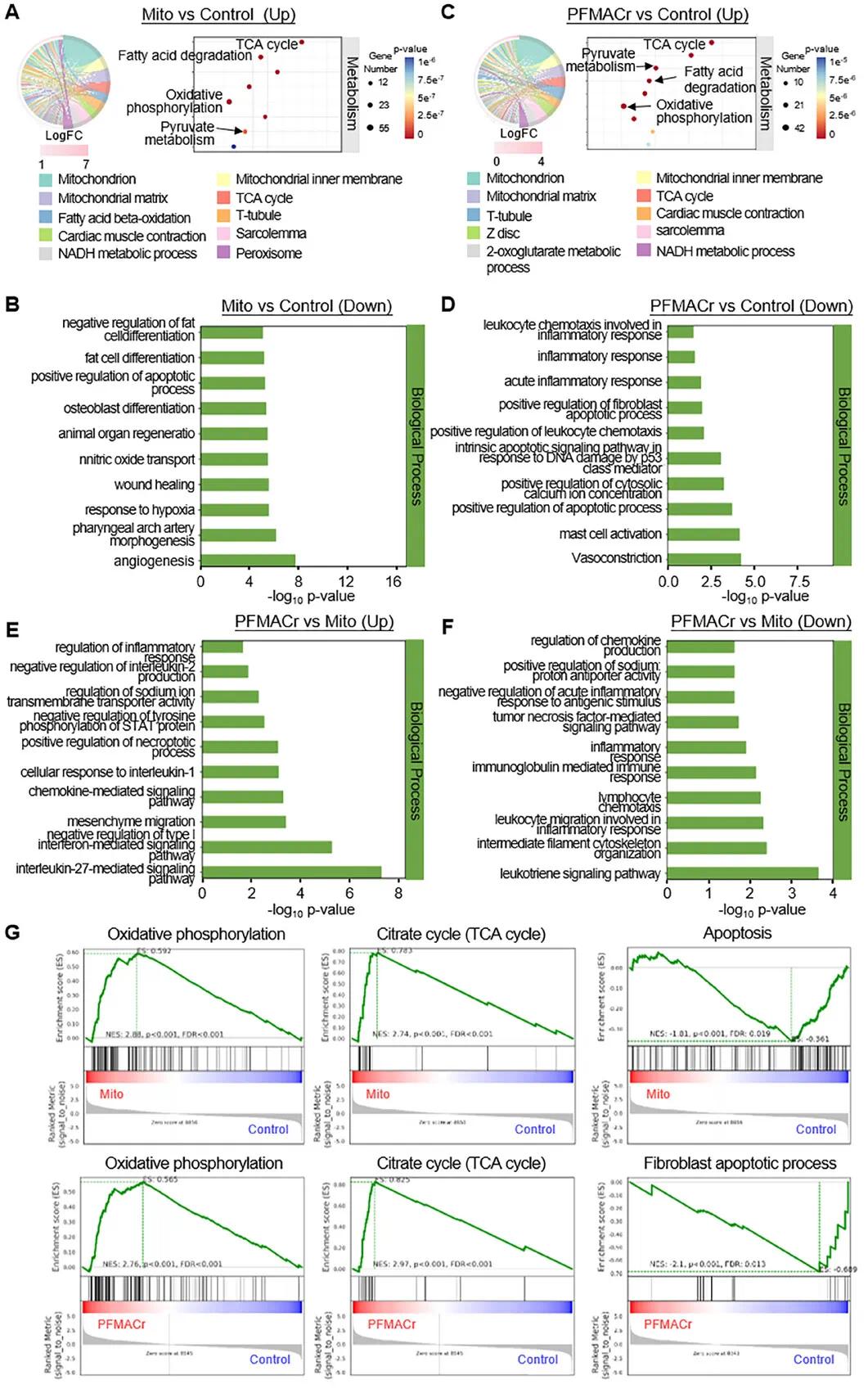
图6 天然线粒体和PFMACr AMN处理的IHD大鼠的心脏转录组学。(a-f)GO和KEGG富集分析显示,与对照组相比,天然线粒体(Mito)组(a,b)和PFMACr组(c,d)中差异表达的基因,以及与Mito组相比,PFMACr组中差异表达的基因(e,f)。弦图:右侧反映分类组成,中间线表示分类和基因对应关系,左边圆圈为GO术语中的DEG;气泡图:气泡大小表示差异表达基因数量;直方图:GO条目名称显示在左侧。(g)应用GSEA比较Mito组和对照组之间以及PFMACr组和对照组之间参与氧化磷酸化、TCA循环和凋亡途径的基因组
该研究构建携带高能磷酸键的人工线粒体纳米机器人(AMNs)重建体内能量供应系统以提供能量。特别是,通过引入屏障穿越模型,这类 AMN 也可以口服给药,为慢性疾病的治疗提供了一种更符合临床的递送方法。 AMN 在胃肠道中保持稳定,通过穿过肠屏障,并使用运动单元以趋化作用靶向受损的心脏组织和心肌细胞。在细胞内,它们的能量产生为 ATP 合成提供高能磷酸键(持续时间 12 小时),同时协同减少炎症和恢复细胞活力。在相同的给药频率下,口服 AMNs(50 mg kg−1)的治疗效果与静脉注射 AMNs(10 mg kg−1)相当,为改善心脏功能提供了一种方便的方法。 在转录组学分析进一步揭示,口服AMNs 不仅上调与 ATP 产生直接相关的基因,恢复受损心脏组织中线粒体的代谢和结构功能,而且通过降低炎症水平和减少凋亡和纤维化过程来修复受损心脏,为通过体内能量传递和病理微环境调控治疗线粒体功能障碍性疾病提供了可行的途径。

|
创赛生物 提供高品质的医疗产品和服务 |
联系我们 |
产品中心 |
扫码关注
关注公众号 扫码加客服
|

